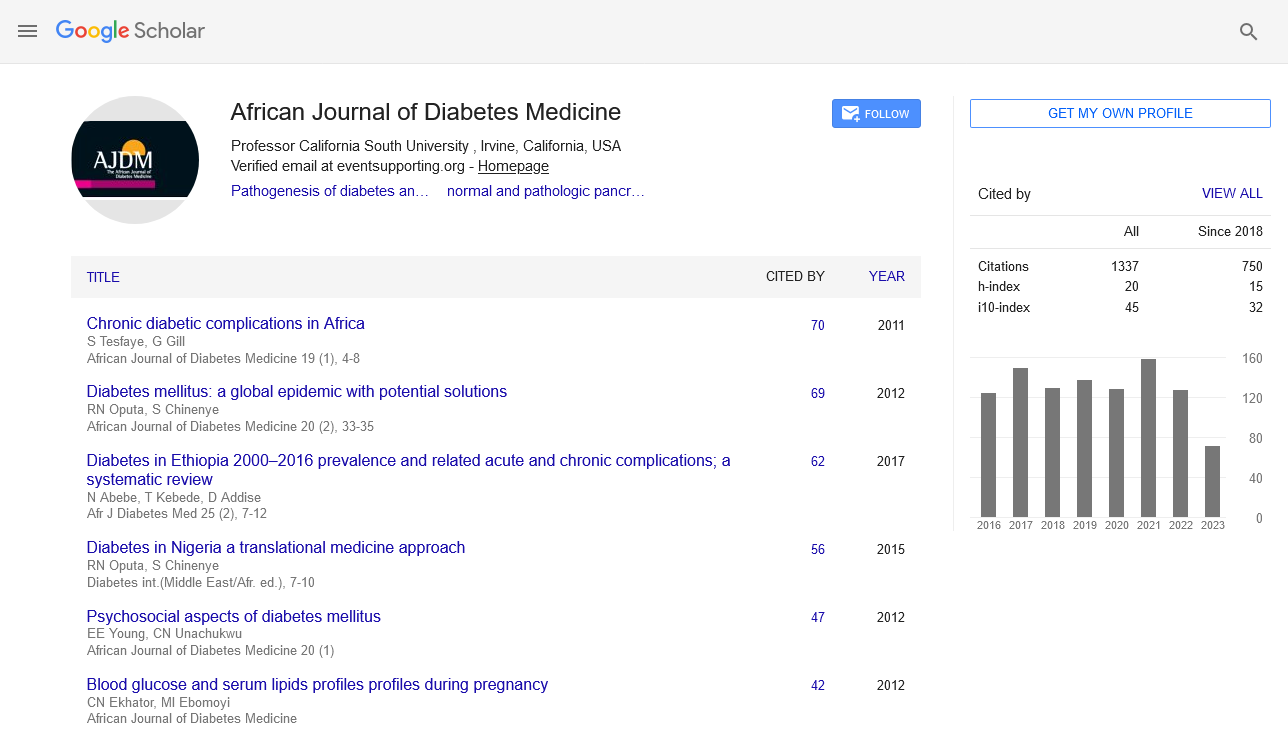
African Journal of Diabetes medicine received 1471 citations as per google scholar report
Select your language of interest to view the total content in your interested language
To read the issue click on a cover